
A dieta cetogênica se distingue por seu baixo teor de carboidratos (~ 5% da ingestão calórica total), proteína moderada (~ 15% da ingestão calórica total) e alto teor de gordura (~ 80% da ingestão calórica total). A adaptação mais comum dessa dieta envolve uma proporção de 4:1 de gordura para carboidratos e proteínas.
Quando a ingestão de carboidratos é insuficiente, a oxidação dos ácidos graxos atinge o pico e a produção de acetil-CoA aumenta nas mitocôndrias dos hepatócitos (células do fígado). Acetil-CoA então entra no ciclo do ácido cítrico (ciclo de Krebs), junto com o oxaloacetato. Quando o oxaloacetato é consumido e sua quantidade não atinge o equilíbrio do ciclo cítrico, o acetil-CoA passa a produzir os corpos cetônicos acetoacetato e β-hidroxibutirato (βHB) como fonte alternativa de energia para os tecidos fora do fígado.
Esses corpos cetônicos podem controlar o uso de substrato, inflamação, estresse oxidativo, processos catabólicos e expressão gênica. Isso é realizado com a ajuda da enzima mitocondrial 3-hidroxi-3-metilglutaril-CoA sintase 2 (HMGCS2) que ativa a primeira etapa da cetogênese. O βHB fornece mais trifosfato de adenosina (ATP) por mol de substrato em comparação com o piruvato.
A cetose é alcançada quando o βHB no sangue atinge concentrações ≥ 0,5 mmol/L. A enzima metabólica succinil-CoA:3-cetoácido CoA transferase (SCOT) não está presente no fígado, portanto, as células do fígado não podem usar corpos cetônicos para obter energia. Assim, o acetoacetato e o βHB podem deixar o fígado e entrar na corrente sanguínea para serem distribuídos para diferentes corpos tecidos, incluindo o cérebro.
A enzima βHB desidrogenase pode converter os corpos cetônicos entre si e essa conversão reduz o dinucleotídeo de nicotinamida adenina (NAD+) a NAD+ (NADH). As cetonas poderiam então ser usadas pelos neurônios do cérebro ou outros tecidos do corpo para reformar o acetil-CoA, que então entra no ciclo do ácido cítrico novamente para gerar ATP.

Corpos cetônicos beneficiam pacientes com câncer
A maior parte dos tumores não pode fazer uso de corpos cetônicos porque não expressam uma ou mais das enzimas β-hidroxibutirato desidrogenase (β-OHBDH) ou SCOT. Assim, o corte dos carboidratos e o aumento da gordura na dieta, gera mais corpos cetônicos e priva as células tumorais energia de que precisam para progressão e sobrevivência.
O potencial antitumoral da dieta cetogênica foi demonstrado em alguns estudos in vitro e in vivo, especialmente nos gliomas, câncer de mama e cancro de cólon. Estes tumores não são capazes de empregar cetonas como fonte de energia dietética quando privadas de glicose, resultando em diminuição da proliferação tumoral. Os corpos cetônicos também melhoram a eficácia de medicamentos usados no tratamento do câncer. Assim, a dieta cetogênica deve ser empregada junto com as terapêuticas tradicionais.
Como os corpos cetônicos reduzem o câncer?
Uma característica do metabolismo do câncer é um aumento na captação de glicose, o que leva a níveis elevados de insulina, que por sua vez eleva o fator de crescimento semelhante à insulina 1 (IGF1). A ligação da insulina e do IGF-1 aos seus receptores tirosina quinase leva à autofosforilação e subsequente ativação da fosfoinositídeo 3-quinase (PI3K).
A PI3K, por sua vez, leva à ativação e autofosforilaçãon da via da proteína quinase B (AKT). A superativação da via PI3K/AKT tem sido associada a níveis baixos de glicose e resulta na morte rápida de células tumorais. AKT estimula o alvo mamífero da rapamicina (mTOR). O mTOR provoca a glicólise aeróbica por meio de sua influência nas principais enzimas glicolíticas, especificamente por meio de seus efetores a jusante c-Myc e fator induzível por hipóxia (HIF)-1a.
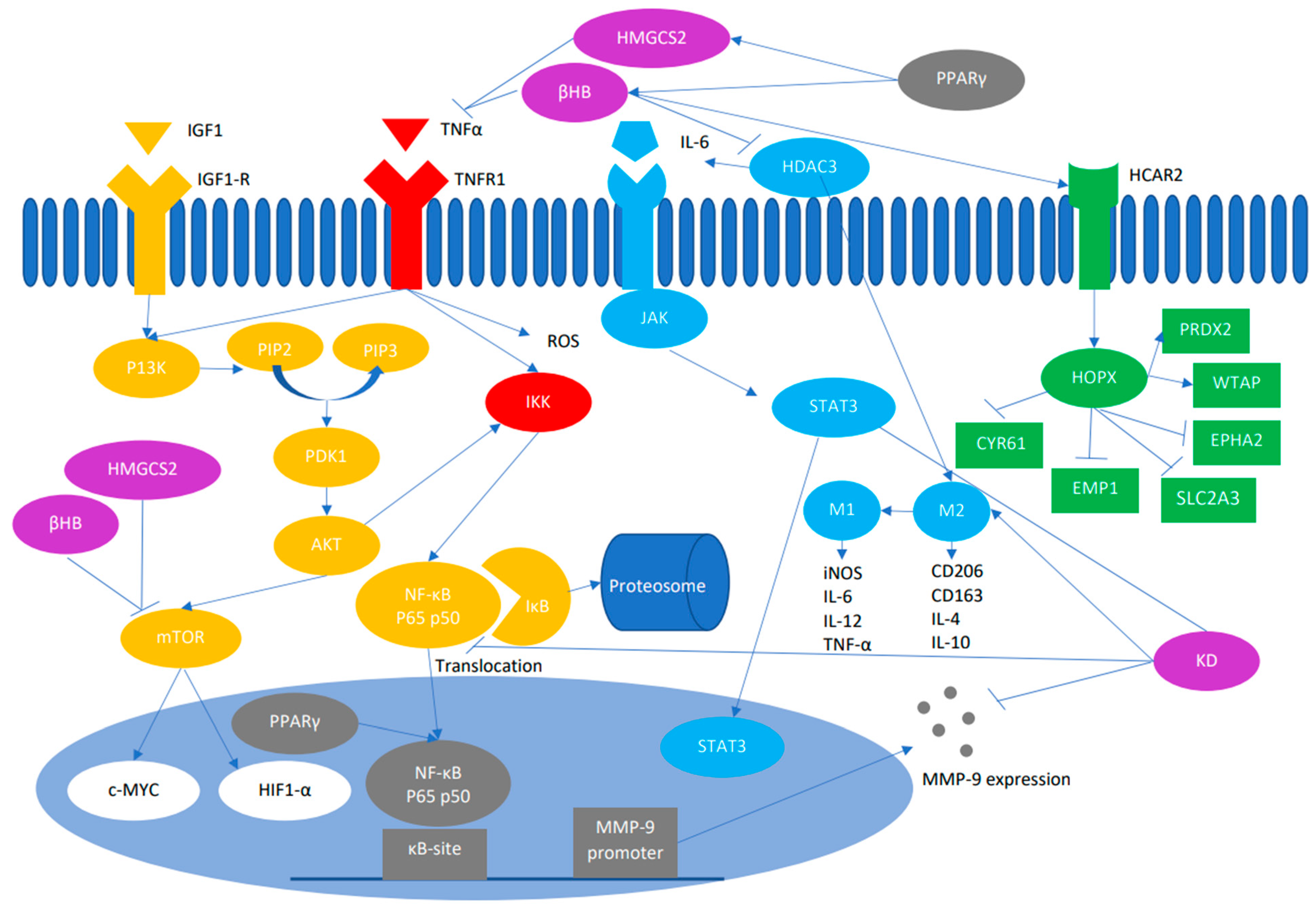
O NF-κB é uma proteína heterodímera composta por duas subunidades, p65 e p50, que são necessárias para ativar e translocar o NF-κB para o núcleo. Estímulos extracelulares, como um receptor do fator de necrose tumoral (em vermelho), interage com seu ligante TNF para regular positivamente o complexo IκB quinase (IKK).
O complexo IKK, por sua vez, fosforila o IκB ligado a p65/p50. O IκB fosforilado é degradado pela via ubiquitina-proteassoma, consequentemente ativando o NF-κB. O NF-κB ativado se transloca para o núcleo, onde ativa a expressão de genes a jusante que possivelmente aumenta a inflamação e a iniciação e progressão do câncer.
A ativação dessa via resulta no aumento da expressão de ciclina D1, ciclina E e quinase dependente de ciclina (CDK)-2, bem como de IL-6 e Myc. A IL-6 ativa transdutores de sinal e ativadores da transdução-3 (STAT3). A sinalização de IL-6/STAT3 tem uma consequência importante nas células imunes que se infiltram no tumor no microambiente imune do tumor no CRC [59].
A histona desacetilase 3 (via azul claro) controla a secreção de IL-6 e participa do aumento do fenótipo protumoral M2 do macrófago. Os macrófagos associados ao tumor (TAMs) são polarizados no M1, que são classicamente ativados e inibem o desenvolvimento do tumor ou no M2, que são alternativamente ativados e aumentam a progressão do tumor e a metástase.
O macrófago de fenótipo M1 aumentou a expressão de moléculas de óxido nítrico sintase induzível (iNOS), IL-12, MHC e CD16/32 e produziu fatores pró-inflamatórios, como IL-6, IL-12 e TNF-α, que pode iniciar a resposta imune. O fenótipo M2, por outro lado, produziu o receptor de Manose C tipo 1 (CD206), Cluster de diferenciações 163 (CD163), arginae-1, IL4 e IL10, que estão envolvidos na resposta anti-inflamatória e cura de feridas.
O NF-κB também está envolvido na expressão da metaloproteinase de matriz (MMP)-9 (Figura 2). A MMP9 é uma proteinase que pode digerir a matriz extracelular e as membranas basais e o colágeno tipo IV subjacente aos vasos sanguíneos, o que facilita a migração de cânceres agressivos.
In vivo, os corpos cetônicos minimizam a expressão da proteína MMP-9 e aumentam a polarização M2 para M1. Além disso, os níveis de proteínas HDAC3/PKM2/NF-κB 65/p-Stat3 diminuem no grupo em cetose.
Corpos cetônicos reduzem efeitos colaterais do tratamento
Oos efeitos colaterais da quimioterapia e da radioterapia podem ser muito incapacitantes e influenciar severamente a qualidade de vida. Alguns desses efeitos colaterais incluem náuseas, vômitos, fadiga, perda de cabelo, danos nos nervos e músculos, comprometimento cognitivo e perda de peso.
Uma dieta baixa em carboidratos e aumento de corpos cetônicos melhoraram a qualidade de vida, controlam os efeitos colaterais da quimioterapia, especificamente náusea e fadiga, e previnem a perda de massa muscular. Mas a qualidade da dieta é importante. Outra opção é suplementar ésteres de cetonas e triglicerídeos de cadeia média, capazes de aumentar corpos cetônicos no sangue, mesmo sem a restrição severa de carboidratos (Tamraz, Ghossaini, & Temraz, 2023).
