
A microbiota intestinal contribui para a regulação da homeostase energética e tem sido associada tanto ao excesso de peso corporal e acúmulo de massa gorda (sobrepeso, obesidade) quanto à perda de peso, fraqueza, atrofia muscular e depleção de gordura (ou seja, caquexia).
A figura abaixo traz as principais áreas cerebrais envolvidas na alimentação homeostática (núcleo arqueado, núcleo paraventricular e hipotálamo lateral) e hedônica (área tegmental ventral, núcleo accumbens, estriado e córtex pré-frontal).

ARC (núcleo arqueado), PVN (núcleo paraventricular), LH (hipotálamo lateral), VTA (área tegmental ventral), NA (núcleo accumbens), estriado e PFC (córtex pré-frontal) - Hul et al., 2024
Efeito dos microorganismos e metabólitos intestinais no contexto da obesidade e comorbidades relacionadas
As bactérias intestinais podem se transformar, decompor e produzir compostos, resultando em uma infinidade de metabólitos microbianos, como ácidos graxos de cadeia curta (AGCC), ácidos biliares (AB), lipídios bioativos, aminoácidos (AA), indóis e vitaminas. Acredita-se que o microbioma humano produza ou processe mais de 55.000 compostos diferentes, muitos dos quais podem modular a fisiologia e a fisiopatologia do hospedeiro de alguma forma. Para tanto, as bactérias usam substâncias prebióticas, incluindo polissacarídeos não-amiláceos, amido resistente, oligossacarídeos resistentes (XOS, GOS, FOS, POS, dextrinas resistentes) e polifenóis.

Fibras prebióticas alteram a composição e/ou a atividade da microbiota intestinal, com produção de AGCC e outros metabólitos que são usados no cólon ou no fígado, afetando suas funções. Metabólitos podem se ligar e ativar receptores acoplados à proteína G, levando à produção de hormônios intestinais, regulando beneficamente a homeostase energética e a função da barreira intestinal.

Funções e efeitos das enterosinas (moléculas derivadas no intestino) e que conseguem modular neurônios entéricos, diminuindo a motilidade duodenal para (i) reduzir a absorção de glicose e (ii) restaurar o eixo intestino-cérebro-periferia e a sensibilidade à insulina durante o dia (Hul et al., 2024)
Nutrição de nova geração: integrando o microbioma nas recomendações alimentares
O microbioma — o conjunto de micro‑organismos que habitam o intestino — funciona como um mediador importante da forma como a alimentação afeta o organismo: metabolismo, imunidade, integridade intestinal, entre outros.

A dieta fornece substratos (por exemplo fibras alimentares, polifenóis, outros micronutrientes) que são metabolizados pelo microbioma, produzindo metabólitos (como ácidos gordos de cadeia curta) que têm efeitos fisiológicos no hospedeiro.

O microbioma, por sua vez, modula digestão, absorção, imunidade, homeostase metabólica e barreira intestinal — o que significa que a “qualidade” da microbiota pode influenciar como o indivíduo responde à alimentação. Assim, as recomendações dietéticas devem levar em consideração a ciência do microbioma.
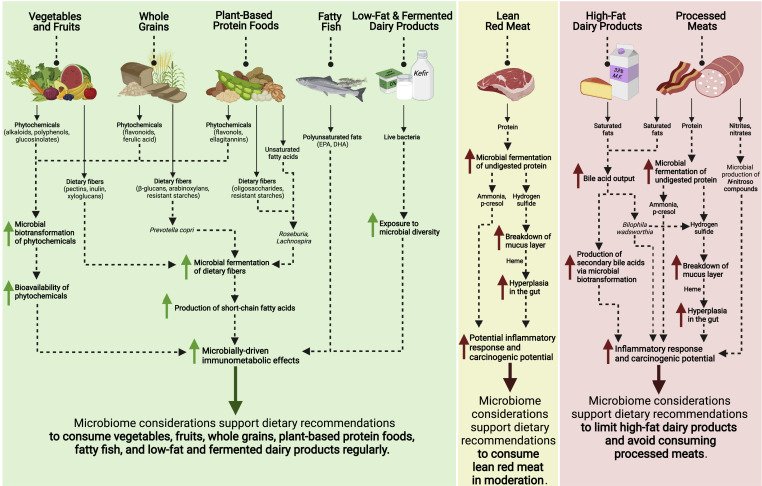
O que comemos, como comemos, com quem comemos, tudo importa. Além de vegetais ricos em fibras e polifenóis, devemos levar em conta o contexto individual (histórico alimentar, genética, estilo de vida) e o microbioma próprio que tornam cada pessoa um “ecosistema” distinto.
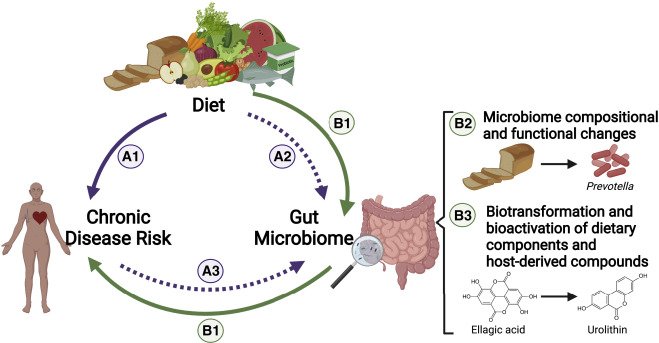
O microbioma e a dieta não são respostas mágicas a tudo, mas sem eles funcionando bem, a saúde é sim afetada. Lembrar que existem outros fatores (genética, ambiente, estilo de vida) que devem ser levados em consideração.
Modulação intestinal (curso online): https://bit.ly/disbiose-at














